第二章.視覺系統解剖與生理(一)
視覺是知覺心理學領域中,研究最廣泛的議題,畢竟人的知覺經驗幾乎超過半數來自於視覺。發揮視覺功能的第一站是眼球,它不僅是『靈魂之窗』,透過它還可以讓我們保持平衡感,而且影響到我們的性徵發育。所以在這一章中我們要分別詳談眼球的保護組織、運動組織與解剖構造與其他視覺生理特性和視覺光學知識,以便有充分的知識理解高層次的視知覺現象。
眼球外部保護與運動組織
脊椎動物的眼球多半是被藏在頭部的前方(例如人眼)或側方(如兔眼)顱骨內。在前方者對深度訊息敏感,但視野便比較小。在側方者視野加寬,但對深度訊息就失去敏感性(詳細狀況將在深度知覺章節中敘述)。因為眼球本身相當脆弱,所以它不只深陷眼框(orbit)內,受到頭骨保護,其周圍尚有許多脂肪(orbital fat)襯墊著,以防強烈震動,前方又有眼臉(eyelids)可以隨時隨意或不隨意的覆蓋,例如眨眼(blink),以防眼球乾澀、或異物碰觸等。其中眼臉的位置可能還透露出這個人對外界事務的興趣高低,例如當他對你說話的內容有興趣時,他可能張大眼睛看著你。
眨眼現象則更有趣。一般眨眼速率約每秒4次,有些時候是不自主的眨眼,但也可以隨意自主的眨眼。當空氣太乾燥、情緒激動時或是用腦筋思考、對談時,眨眼速率多會增加(Doane,1980)。每一次眨眼所需時間約1/3秒,這1/3秒內有99%的外界光線被擋掉,但是我們並不會覺得外界突然變得昏暗,為什麼呢?美國布朗大學的心理學家Volkmann、Riggs與Moore (1980)做了一個很漂亮的實驗解決了這個迷團。他們在暗室內用一條透明的彈性光纖將強光經由嘴巴內的上顎直接刺激到眼球網膜(注意,即使眼皮蓋住仍能『感覺到』該強光),然後透過微電極偵測受試者在眨眼期間,對該強光的亮度偵測閾值與非眨眼期間對該強光的亮度偵測閾值。結果發現眨眼期間受試者對強光的偵測閾值顯著高於非眨眼期間。由於該強光並不會因眨眼而被遮蓋住,因此推論:眨眼時的亮度偵測閾值增加是因為,視覺系統在眨眼期間也暫時關閉所致!
至於人眼的運動則主要是由六條肌肉(extraocular muscles)協同牽動。其詳細運作機制將在運動視覺一章中詳述。
眼球的內部解剖結構
眼球壁的結構基本上可以分三層:角膜鞏膜層(corneo-scleral layer)、葡萄膜層(Uveal layer)及視網膜層(retinal layer)。
角膜鞏膜層包括鞏膜(sclera)與角膜(cornea)。鞏膜是白色不透光的結締組織,佔約整層5/6的面積,它讓六條眼外肌(extra-occular muscle)的終點附著;角膜的曲率比鞏膜大,佔1/6面積,是透明的,可吸收紫外線,其組織纖維不是靠血液吸收養分,而是靠anterior chamber內的液體供給養分,所以它幾乎是透明的,也是光線產生折射(refraction)的第一道關卡。它與鞏摩接合處有個凹陷,稱limbus。anterior chamber則內含玻璃液(aqueous humor),除了提供角膜與水晶體養分外,也可維持眼球形狀。
葡萄膜層包括脈絡膜(Choroid)、睫狀體(ciliary body)、虹膜(iris)與血管組織。脈絡膜有很深的黑色素可吸收穿過網膜的多餘光線,並提供養分給網膜層;睫狀體是指由一塊平滑肌與懸韌帶(suspensory ligment or zounle)組成,可把水晶體(lens)架起,並調節水晶體厚薄的組織;虹膜則是一片含有環狀排列與輻射狀排列的平滑肌,蓋在水晶體前緣,以形成瞳孔(pupil)調節光線進入的量。從角膜到水晶體所構成的光線折射界面,形成複雜的光學系統,這將在最後一節中詳細介紹。
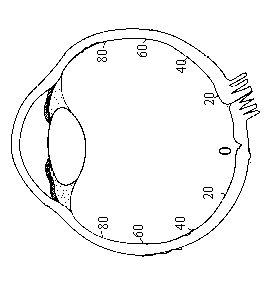
視網膜層是眼球壁最內層,唯一含感光細胞與神經細胞的構造。基本上又可分色素層(pigment layer)與神經層(neural layer,在解剖上又分出9層,此處無必要如此詳述)。色素層只是一層色素上皮細胞(pigment epithelial cell)組成,與脈絡膜相隔一層Bruch
’s膜。接下來的神經層由外向內依序有感光細胞,包括錐細胞(cone)與桿細胞(rod,參見圖2-5),其構造容後再詳述。還有水平細胞(horizontal cell,主要是一種抑制性細胞)、雙極細胞(bipolar cell,一端與光受器連接,另一端與節細胞或無軸索細胞連接。從型態來分,有的雙極細胞很小,只連接一個光受器,被稱為midget bipolars;大多數則可連接多個光受器,被稱為diffuse bipolars)、無軸索細胞(Amacrine cell,主要與雙極細胞和節細胞產生突觸,可將雙極細胞的訊號水平擴散到好幾個節細胞上,也可將數個雙極細胞的訊息集中到一個節細胞上)、節細胞(ganglion cell)。當錐細胞直接傳遞給雙極細胞興奮訊息,則藉著水平細胞傳遞抑制訊息,反之,當錐細胞直接傳遞給雙極細胞抑制訊息時,則藉著水平細胞傳遞興奮訊息(參見圖2-8)。整個視網膜的中央部位有一個無血管區域,就稱為黃斑區(macula,因有黃色素而命名),直徑約1-3mm,而黃斑區中央最薄(之所以最薄是因為此處的節細胞等組織幾乎全被推擠到旁邊)的地方稱中央小窩(fovea),約佔1-2度視角(以中央小窩為0度視角起算點,1度視角也稱為1度離心度,即eccentricity)。雖然中央小窩只佔不到2度視角,但卻是視覺最最重要的區域。一般而言,在中央小窩處的錐細胞與雙極細胞幾乎是一對一直接連結,但桿細胞藉著水平細胞與雙極細胞的連結,則高達數十個、數百個。
不管是錐細胞或桿細胞,其結構都可分三層,包括外段(outer segment)、內段(inner segment)與纖維(fiber,分別稱為rod fiber與cone fiber)。其中,纖維的主要功能在與其他細胞形成突觸,內段含粒腺體、內質網等胞器。只有外段會對光刺激反應,因為外段含有視紫素(visual purple,或稱rhodopsin,蓋因此色素是從Rod中抽取出來,故名之)。該視紫素又是由一種叫opsin的蛋白質與胡蘿蔔素(retinal,即vitamin A aldehyde)共同組成(Wald,1964),在完全黑暗的情況下,感光細胞(包括rod與cone)會不斷釋放興奮性的神經傳導物質,不過,當光刺激它時,視紫素將產生一連串生化反應,造成過極化(hyperpolarization)而停止釋放神經傳導物質。
這些色素的反應只會隨著打進來的光子(光子可以想像成光波的最小能量單位)數目增加而增加,而不論這個光子本質是短波或長波,此現象被稱為等變異原則(principle of univariance,Rushton,1972)。例如
錐細胞與桿細胞的差異基本上有3項:(1)錐細胞(cone)的視紫素可分三類,對不同波長的光能有不同吸收反應,其最高反應所對應的波長分別約是420、530、560nm(Bommaker,Dartnall與Mollon,1980),因此我們慣稱這三類錐細胞分別為S Cone、M Cone、L Cone;(2)兩者在網膜上的分佈相當不平均,桿細胞由ora serrata向中央小窩(fovea)逐漸減少,錐細胞則多在中央小窩中,中央小窩 2度視角內幾乎無桿細胞,而且錐細胞成六角形碼賽克(mosaic)排列;(3)桿細胞個數約有1.3x108,錐細胞只有約7x106。
節細胞又稱optic tract neuron,其軸突從網膜各個方向往視乳頭(optic papillae,或稱optic disk)集中,然後穿過鞏膜,形成一束視神經。該束視神經軸突在網膜內所『鑿』開的區域約佔5度視角。該區域為了讓視神經軸突通過,而無任何感光細胞存在,故又稱盲點區(blind spot),意即吾人單眼的視野中,有一塊佔5度視角的區域是看不到的,讀者可以試著依照圖2-9的指示,閉上一眼,用另一眼(單眼)觀看以感覺一下自己盲點區域存在的事實。
節細胞是整個視覺通路上第一個在解剖與生理上有清楚樹突、軸突區分的細胞,而且其軸突在網膜內沒有包上髓鞘,出了網膜就被包上髓鞘了。
在型態上,節細胞又分兩類:一類細胞本體比較大,樹突多的,稱為alpha節細胞;另一類細胞本體小,樹突小的,稱beta節細胞(Boycott & Wassle,1974)。在功能上分類,則可分X節細胞,是對於光亮或光暗之後才逐漸產生持續活動電位反應的節細胞;Y節細胞,則是在光亮或光暗一剎那產生短暫的(transient)活動電位反應(Enroth-Cugell與Robson,1966)。到目前為止尚未清楚,從功能分類出的X、Y節細胞與從型態上分類出的alpha、beta節細胞之間存在什麼樣的對應關係。
節細胞總共約106 個,大約只有全部感光細胞數目的1/120,可見它對於感光細胞所承載的訊息做了強力壓縮!但壓縮比率不均勻,網膜週邊壓縮比率比fovea區壓縮比率大,它們的功能將在後面章節中詳述。
視神經到視皮質區的路徑
網膜上的節細胞軸突在視乳頭處離開眼球之後,隨即被oligodendroglia包住,形成有髓鞘的神經纖維,而且左右兩眼的視神經在視交叉處(optic chiasm(a))會合。之所以稱為視交叉是因為,左右兩眼的視神經纖維在此處會合之後,負責左眼的左視野(即將視網膜分成左右兩半之後的鼻側部份)與右眼的左視野(即視網膜之顳側部份)視神經共同形成右側的optic tract;反之,左眼的右視野(即視網膜之的顳側部份)與右眼的右視野(即視網膜之鼻側部份)視神經共同形成左側的optic tract。這種左右視野訊息交叉投射到右左腦的現象與各種不同視野缺損的病症有密切關連。
到目前為止,從節細胞以後(包括節細胞)的神經細胞才開始有真正的活動電位,而且節細胞本身也有自發性的活動電位產生,這類自發性神經衝動對視覺感受而言,扮演極重要的角色。
在視交叉會合的視神經大部分形成左右optic tract之後,分別投射到左右側膝核(lateral geniculate nucleus,以下簡稱LGN)。注意,這種投射是點對點的投射,如圖2-12所示,網膜的a、b、c、d區各對應到側膝核的a、b、c、d區,又稱為網膜拓樸對應(retinotopic map)。
LGN被染色後可分出6層型態不同的細胞,其中,第2、3、5層接受同側眼(ipsilateral eye) 顳側(temporal side)之網膜節細胞訊息輸入,第1、4、6層接受異側眼(contralateral eye)鼻側(nasal side)之網膜節細胞訊息輸入。意即左側LGN的第2、3、5層接受左眼的右視野網膜(即顳側網膜)訊息,第1、4、6層接受右眼的右視野網膜(即鼻側網膜)訊息;右側LGN的第2、3、5層接受右眼的左視野網膜訊息,第1、4、6層接受左眼的左視野網膜訊息。左右兩眼的訊息雖可能在同一側LGN內相遇,但是在各層間的左右眼訊息幾乎無任何交互作用(R.L. DeValois,1965),即LGN的每一個分層都對應整個眼球的半視野,層與層之間極少產生交互作用。
而且根據Leventhal、Rodieck & Dreher(1981)的研究,alpha型節細胞主要多投射到LGN的第1、2層,形成樹突長而疏,細胞大,傳導快,反應持續短的magnocellular層(以下簡稱magno層);也有一小部份alpha型節細胞投射到parvocellular層(以下簡稱parvo層)。beta型節細胞則投射到第3、4、5、6層,形成樹突短而密,細胞小,傳導慢,反應持續長的parvo層。LGN不只是接受網膜投射的訊息,也接受腦幹(brain stem)、視丘(thalamus)、其他視皮質區的回饋(feedback)投射(J.S.Lund,R.D.Lund,Hendrickson,Bunt與Fuchs,1975)。
Peter Schiller、Nikos Logothetis與Eliot Charles(1990)曾將猴子(rhesus monkeys)的LGN分層破壞(注射神經毒素ibotenic acid),觀察猴子原先學會的形狀、顏色、運動、深度、質地的辨識行為(被條件化的行為)是否受到影響,結果發現magno層的破壞使得猴子無法偵測運動,parvo層破壞則使它無法偵測色彩、細微質地、形狀、深度辨識。我們可以粗略地整理如下的比較表:
|
Parvo層 |
Magno層 |
|
|
解剖上的差異 |
細胞體小 樹突細密 數目多(尤其是fovea區)
|
細胞體大 樹突疏長 數目少(多在para-fovea區) |
|
生理反應上的差異 |
傳導慢(20m/sec.) 反應持續長 R.F.小 對顏色敏感 對比敏感度低 同樣illumination pattern旋轉不反應 |
傳導快(40m/sec) 反應持續短 R.F.大 對顏色不敏感 對比敏感度高 同樣illumination pattern旋轉反應 |
|
對應功能上的可能差異 |
形狀細節辨認 空間分析 色彩處理 深度偵測 |
偵測運動 時序分析
|
但是請注意,有一部分(約15-20%,多是M型節細胞)視交叉會合的視神經沒有投射到LGN,而是投射到上小丘(Superior Colliculus),這個路徑被稱為tectopulvinar視覺系統,以別於從視交叉、LGN到視皮質區的geniculostriate系統。投射到上小丘的系統可能與眼睛的光反射(包括強光時瞳孔縮小、雙眼注視遠近物體時的水晶體調節、兩眼輻輳動作,以及平常看東西時,頭不自主的向前傾、舉起手臂檔光或保護眼睛等視覺性的身體反射動作)有關(Wurtz與Goldberg,1971)。上小丘的解剖及生理功能頗為複雜,因為上小丘不只接受側膝核的訊息,也接受V1回饋的訊息,對其他感官通道(sensory modality)傳來的刺激也有反應,我們不打算詳述它。
由LGN投射到大腦初級視皮質區(以下簡稱V1)的纖維因呈現輻射狀,故又稱視輻射(optic radiation)。視輻射的神經纖維基本上有兩個路徑,一個繞過側腦室(lateral ventricle),另一個繞過側腦室的顳角(temporal horn),終點都在禽距溝(calcarine sulcus)上下的腦皮質區域。依Brodmann的分類法,該皮質區屬於第17區,因為有清楚的帶狀紋,故又稱橫紋區(striate cortex)。而這塊區域與側膝核也有點對點的投射關係,稱之為拓樸對應(topographical mapping)。重要的是,網膜上小小的黃斑區透過側膝核所對應的皮質區就佔了1/3。
Perry & Cowey(1985)找到生理證據顯示:從網膜中央投射到側膝核的P系統有些微放大投映現象,從側膝核到視皮質區也有放大,總共約放大25%的訊息,此現象稱為皮質放大效應(cortical magnification)。
Hartline(1940)定義:能夠影響某一特定神經元被興奮或抑制的空間範圍或與此特定神經元產生輸入連結的感覺受器(receptor)範圍,就被稱為該一特定神經元的接受域(receptive field,以下簡稱R.F.)。如前述雙極細胞與感光受器的神經網路連結方式來看,我們可以想像雙極細胞對其R.F.範圍內的不同空間位置之光刺激的反應型態如下圖 所示。
美國MIT教授David Marr(1980)以兩個極性相反、不同標準差的常態分配曲面之加成函數來模擬如圖 的R.F.反應型態,並稱這個加成函數為DOG(Difference Of Gaussian)函數。
我們發現1個節細胞接受數十個感光細胞的訊息,1個側膝核細胞又接受數個節細胞的訊息輸入, 1個V1細胞又接受數個側膝核細胞的訊息輸入,則越後站的細胞的R.F.理論上應該越大,似乎訊息傳到越後面,應該壓縮得越厲害,實際上R.F.仍有變小的時候。而且從網膜到側膝核雖然發生訊息壓縮,但是總體而言,網膜中央小窩(fovea)的訊息到了皮質區仍是放大的。
Hubel & Wiesel(1961)比較側膝核與節細胞的接受域,結果發現它們都非常相似,都是呈現圓形,有內興奮外抑制(on-center off-surround)或外興奮內抑制(off-center on surround)兩型。為什麼它們會有這類接受域反應型態?讀者可從圖 與圖 中看出端倪。
幾乎所有初級皮質區在解剖上都約略可分出6層,如圖 。由側膝核投射到V1的纖維首先停在中間的第4層。其中magno細胞的突觸端停在4cα(V1的第4層又分a、b、c三層,4c層又分α、β兩層),parvo則停在4cβ 或4a層。
1970年之前許多研究者多以為橫紋區之外的視覺皮質區(又稱外橫紋區,extrastriate cortex)只是扮演視覺記憶的角色,但自從1970年代,許多神經生理學家開始在這些外橫紋區上找到對不同視覺刺激特徵反應的細胞之後,這個想法就改變了。他們認為橫紋區之外仍有不同的、平行的視覺功能管道。Leslie Ungerleider與Mortimer Mishkin(1982)在猴腦上做手術,觀察到若將頂葉(parietal lobe)毀除,並不影響猴子做物體區辨作業(object discrimination task,即在兩個形狀不同的物體中,選出一個剛剛出現過的、熟悉的物體),但卻使得該猴子無法做地標區辨作業(landmark discrimination task,即在一個有圓柱做標記的位置旁放有布遮蔽了的食物,另一個比較遠的位置只有布沒有食物,猴子必須學習認出食物只有在圓柱標記旁而已)。反之,若將顳葉(temporal lobe)毀除,則不影響猴子做地標區辨作業,只影響該猴子無法做物體區辨作業。這個功能雙分離(double dissociation)的證據強烈顯示,在外橫紋區必有兩個路徑分別涉及辨認物體是什麼(what)以及辨認物體在哪裡(where)兩類視覺功能。後來經由許多生理學家的追蹤,大致可簡略畫出這兩條路徑如下:
magno系統經由4c往4b層、V2、V3到中顳葉皮質區背側(dorsal Medial Temporal cortex),再到頂葉,這條路徑涉及物體在哪裡(where pathway)。parvo系統則由4c往2、3層到V2、V4,再到下顳葉皮質區(Inferotemporal cortex),這條路徑涉及物體是什麼(what pathway)。如此分開來敘述,並不代表神經傳導路徑之間有很清楚的界限,只是在功能上它們似乎攜帶不同的視覺訊息,扮演不同的角色而已。事實上V1之後的路徑多有相互連結(reciprocal connection),其連結之複雜度可見圖 所示。
不只是上述傳導路徑的找尋,連帶不同區域的視神經細胞所偏好的反應刺激型態也有很多有趣的發現。最重要的發現莫過於David Hubel與Torsten Wiesel(1950)在貓的視覺橫紋區所找到的柱狀組織(columnar organization),他們因為這個發現而得1981年諾貝爾生理醫學獎。他們在貓腦中插上微電極以記錄該區域細胞的神經衝動訊號,然後在其視野中隨機打出光點,以尋找該細胞所對應的接受域(R.F.)。經過一番隨機嘗試後,他們發現有些區域的細胞對光點反應,但有些細胞對光棒(light bar)反應,還有對不同方向的光條有特別大的反應,總整理一下大約可分出三大類細胞:簡單細胞(simple cell)、複雜細胞(complex cell)與端點細胞(end-stopped cell)。
簡單細胞有長條狀的R.F.(參閱圖 ),中央是興奮區,周圍是抑制區,意即它們對長條的光棒反應最強(也對靜態的小光點有反應,只是不強),而光棒在興奮區出現時,會使該神經衝動速率迅速增加,在抑制區出現則使神經衝動速率顯著減少!當然,如果光棒方向與R.F.垂直時,該細胞神經衝動幾乎消失。因此它是方向性敏感的細胞,我們可以畫出其方向敏感度曲線(orientation tuning curve)如圖 。
複雜細胞除了對特殊方向的光棒有反應之外,對於光棒朝某個方位(direction)移動時,反應更明顯!(參見圖 )。端點細胞(又稱hypercomplex cell)則是對特定長度的線段或角的移動敏感。
更重要的是,當Hubel與Wiesel有系統的改變電極探針的深度與方向,從三個互相垂直的方向,漸漸插深入皮質區內,測量不同區域的細胞反應時,發現這些細胞的反應與R.F.的變化也有系統性的漸變。首先是對應的R.F.有系統的隨斜插深度相鄰排列,被稱為位置柱列(location columns),其次是沿著灰質區表面相鄰的不同部位插入,可測得順時鐘方向漸變的方向敏感的細胞柱列,此稱為orientation columns;再從另一個方向斜插,可測得只對某一眼刺激較敏感的細胞群,左右眼輪流交疊,形成所謂的ocular dominance columns。這三類柱列都大約每隔1mm寬度,重複一次共同的細胞群表徵,Hubel與Wiesel很自然將它們看成一個個處理模組(processing module),並稱之為hypercolumn,如圖 所示。
讓我們再回頭看看,節細胞的R.F.又是長成什麼樣子?它是中央與周圍功能拮抗的(center-surround antagonism)圓形模樣,如下圖 。節細胞的R.F.之所以長成這個形狀,可能是因為因個節細胞要接受數個感光細胞的訊息時,排列在R.F.中央的感光細胞與排列在R.F.周圍的感光細胞各送出相反的神經衝動訊號所致,例如一群是送出抑制訊號,另一群則送出興奮訊號。
這些神經網路的連結方式其實決定了吾人的日常視覺感受!後一章節將會再度詳述。